مقدمه
اهمیت اسیدهای چرب غیر اشباع لینولئیک و لینولنیک در یک مجموعه مقالات از اواخر دهه 1920 تا اوایل دهه ی 1930 بررسی شدند (Burr et al., 1932). این ترکیبات در ساختمان غشاء سلول شرکت میکنند و علاوه بر این پیش ساز اسیدهای چرب دیگری نیز هستند که در تنظیم سوخت و ساز بدن و در عملکرد غشای سلول نقش کلیدی دارند و در حیات تمامی پستانداران ضروری میباشند.
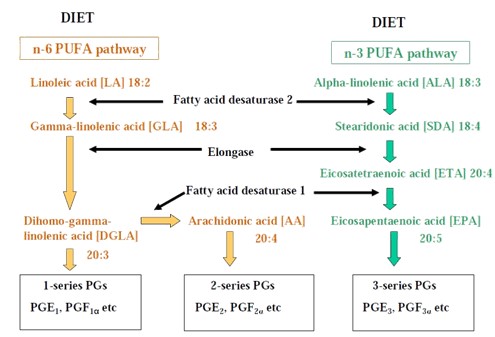
اسیدهای چرب ضروری با محل و تعداد پیوندهای دوگانه سیس در ساختمان مولکولی شناخته می شوند. هر پیوند دوگانه سیس نشان دهنده ی یک خمیدگی 120 درجهای در داخل زنجیره کربنی می باشد، بنابراین با افزایش تعداد پیوندهای دوگانه، زنجیره شکل مارپیچ به خود می گیرد. زمانی که زنجیره های اسیدهای چرب در داخل ساختمان غشاء به کار برده می شوند، زنجیره مستقیم اسیدهای چرب اشباع به طور تنگاتنگی کنار هم قرار می گیرند این در حالی است که اسیدهای چرب غیر اشباع به طور ناپایدار کنار هم قرار می گیرند. در این حالت هرچه باز شدگی لایه ها بیشتر باشد نقطه ذوب کاهش و غشاءها انعطاف پذیرتر می شوند. ساختارها و مسیرهای تبدیل اسیدهای لینولئیک و لینولنیک به محصولات اسیدهای چرب بلند زنجیر در شکل 1 نشان داده شده است. هر چند میبایستی در نظر داشت که فرآیند طویل سازی و غیر اشباع سازی محصولات (که قابلیت طویل شدن دارند) از اسیدهای لینولئیک و لینولنیک توسط ∆-6 دسچوراز (Δ-6 desaturase) انجام می شود و بنابراین این اسیدهای چرب بازدارنده های رقابتی برای یکدیگر محسوب می شوند. با این حال میل ترکیبی ∆ -6 دسچوراز (Δ-6 desaturase) به اسیدهای چرب امگا 3 بالاتر است.
همچنین غلظت های بالای هر یک از این دو (یا هردو) اسید چرب و نیز غلظت های بالای اسیدهای چرب طویل روغن ماهی، از طویل شدن بیشتر زنجیره و غیراشباع سازی اسیدهای لینولئیک و لینولنیک در تولید ایکوزانوئیدهای متناظرشان ممانعت میکنند.
فیزیولوژی و متابولیسم
اگرچه ممکن است اسیدهای چرب ضروری در بافت ها به صورت تری آسیل گلیسرول ها ذخیره شوند، اما نقش های کلیدی آنها در متابولیسم، شرکت در فسفولیپیدهای غشاء و تبدیل به اسیدهای چرب بلند زنجیر و ایکوزانوئیدها می باشندکه به عنوان پروستاگلاندین ها شناخته می شوند. در ارتباط با این بحث مختصر، اسید لینولئیک (6-ω) به اسید آراشیدونیک (AA) تبدیل می شود که یک پیش ساز برای پروستاگلاندین های سری دو می باشد، در حالیکه اسید لینولنیک (3-ω) به ایکوزاپنتاانوئیک اسید (EPA) تبدیل می شود که این نیز یک پیش ساز پروستاگلاندین های سری سه محسوب میشود.
اسیدهای چرب ضروری و ایکوزانوئیدها مهمترین تنظیم کننده های عملکردهای فیزیولوژیکی و بیان ژن محسوب می شوند (شکل 2)، و با باند شدن به مولکول های تنظیمی مختلف با ژن ها ارتباط برقرار میکنند. متابولیتهای اسید آراشیدونیک مثل PGE2 و لوکوتراینهای سری 4 (بین دیگر لوکوتراینها) عموماً مسبب پاسخ پیش از التهاب شامل تب، درد، اتساع رگ ها و نفوذ پذیری عروق می شوند در حالیکه متابولیت های ایکوزاپنتاانوئیک اسید دارای اثرات مخالف بوده یا اثرات کمتری روی این فرآیندها دارند.
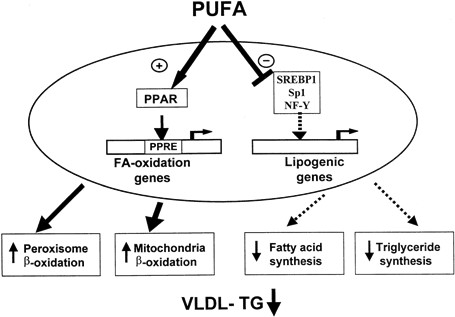
FA: اسیدهای چرب ، NF-Y: فاکتور هسته ای Y، PPAR: رسپتور فعال کننده تکثیر پراکسی زوم ، PPRE: عامل پاسخ فعال کننده تکثیر پراکسی زوم
SP-1: پروتئین القایی 1، SREBP-1: پروتئین باندکننده -1 عامل تنظیم استرول، TG: تری گلیسرید
علائم کمبود و احتیاجات اسیدهای چرب ضروری
گوسالههایی که با جیره های عاری از چربی تغذیه شدند علائم آشکاری از کمبود اسیدهای چرب ضروری را نشان دادند، هرچند کمبودهای خاص نشان داده نشد. این علائم به اختلال در عملکرد غشاءهای سلولی مثل پوشش موی خشن، ریزش مو، خشکی و زبری پوست، آماس پوست، از دست رفتن بیش ازحد آب و … و اختلال در عملکرد اعصاب مربوط است. کمبودها بیشتر مربوط به اثرات ایکوزانوئیدها بر عملکردهای فیزیولوژیکی و تنظیم بیان ژن هستند. هیچ احتیاجی از اسیدهای چرب ضروری برای نشخوارکنندگان مشخص نشده است.
هولمن (1960) تعریفی بیوشیمیایی را برای کمبود اسیدهای چرب ضروری به صورت نسبت اکوزاتری انوئیک به اکزاتتراانوئیک اسید در غشاء گلبول های قرمز بیش از 4/0 گزارش کرد. در شرایطی که اسیدهای چرب ضروری کافی باشد، اسید لینولئیک اسید، ایکوزاتتراانوئیک (آراشیدونیک اسید، 6ω- 20:4) مورد نیاز برای سنتز غشاء سلولی را فراهم میکند. در غیاب اسید لینولئیک کافی، اسید چرب غیر ضروری اولئیک 9-ω غیر اشباع سازی و طویل شده و به شکل اکوزاتری انوئیک اسید (مآد اسید 9ω- 20:3) در آمده که در ساختمان غشاء سلولی شرکت میکند. هولمن (1960) مشاهده کرد زمانی که نسبت این اسیدهای چرب در داخل ساختمان فسفولیپیدهای پلاسما یا غشای گلبول های قرمز موش های جوان از 4/0 تجاوز کند علائم آشکاری ظاهر میشود.
تحقیقات هولمن و همکاران نشان داد که یک اختلاف جنسیتی در میزان نیاز به اسیدهای چرب ضروری وجود دارد. نرهای اختهی مکمل شده با استروژن احتیاجاتی شبیه مادهها داشتند. مطالعات بعدی نشان داد که افزایش فرآیند پاسخ به تحریک تبدیل لینولئیک اسید به DHA در زنان باردار منتسب به اثرات استروژن است (Williams and Burdge, 2006). اینکه آیا این حالات در نشخوارکنندگان و دیگر پستانداران رخ میدهد یا نه ناشناخته است. همچنین پیشنهاد شده که دامنه بهینه برای مصرف اسیدهای لینولئیک و لینولنیک نسبتاً محدود است، بطوریکه در زمانی که مصرف بیش از اندازه است، رشد کاهش مییابد. مصرف بیش از اندازه اسیدهای چرب غیراشباع بدون در نظر گرفتن ویتامین E کافی موجب مایوپاتی شدید ماهیچهای در گوساله ها میشود و مشابه آن در کمبود سلنیوم نیز مشاهده می شود.
گزارش شده است که شیرخوردن نوزاد نشخوارکنندگان تنها چند روز برای جلوگیری از علائم کمبود کافی است (حتی زمانیکه با یک جیره عاری از چربی نگهداری شوند). درحالی که اکسید شدن اسیدهای چرب غیر اشباع در غیر نشخوارکنندگان ارجحیت دارد، نشان دادند که اسید لینولئیک در گوسفند کمتر از 5 درصد از میزان پالمتیک، استئاریک و اولئیک اسید اکسید میشوند. علاوه بر این در زمان گرسنگی، اسید لینولئیک تا حد بسیار کمتر از اسیدهای چرب دیگر بسیج میشوند. 40 تا 50 درصد اسید لینولئیک جذب شده از روده با فسفولیپید لیپوپروتئینهای روده ای ترکیب شده و از اکسیداسیون یا ترشح به داخل شیر حفظ میگردند.
بنابراین با ترکیب شدن بخش قابل توجه از اسید لینولئیک با فسفولیپیدها و استرهای کلستریل (نسبت به اسیدهای چرب غیر استریفیه و تری آسیل گلیسرول) نشخوارکنندگان قادرند، اسیدهای چرب ضروری را بطور بسیار کارآمدی حفظ کنند و با مصرف بسیار کم اسیدهای چرب ضروری نسبت به غیر نشخوارکنندگان زنده بمانند.
احتیاجات به اسیدهای چرب ضروری برای عملکردهای فیزیولوژیکی
رشد
با توجه به آنچه گفته شد، هیچگونه شواهدی که نشخوارکنندگان در حال رشد احتیاج بیشتری برای اسیدهای چرب ضروری فراهم شده از جیره های معمول دارند، وجود ندارد. بر اساس تحقیقات انجام شده، دام های جوان در حال رشد به 88 میلی گرم اسید لینولئیک به ازای هر کیلوگرم وزن متابولیکی بدن نیاز دارند. با استفاده از این مقایسه، مثالهایی از احتیاجات برای اوزان مختلف بدن در جدول 2 خلاصه شدهاند. حتی با توجه به اینکه غشاء سلولی حیوانات بزرگتر (مثلاً نشخوارکنندگان) نسبت به حیوانات کوچکتر (مثلاً موش) دارای سهم بیشتری از اسید لینولئیک است به نظر میرسد که مقادیر موجود بیش از مقدار کافی برای برآورده کردن احتیاجات باشند(جدول 3).
جدول 2 – اسید لینولئیک مورد نیاز برای رشد و نگهداری نشخوارکنندگان، با فرض احتیاج به 88 میلی گرم به ازای هر کیلوگرم وزن متابولیکی بدن.
| وزن بدن (kg) | اندازه متابولیکی بدن (4/3kg) | اسید لینولئیک مورد نیاز (g/day) |
|---|---|---|
| 50 | 18/8 | 1/6 |
| 100 | 31/6 | 2/8 |
| 200 | 53/2 | 4/7 |
| 300 | 72/1 | 6/3 |
| 400 | 89/4 | 7/9 |
| 500 | 105/7 | 9/3 |
| 600 | 121/2 | 10/7 |
شیردهی
برای تعیین احتیاجات شیردهی می بایستی مقدار اسید لینولئیک ترشح شده در شیر مورد بررسی قرار گیرد. فرآیند بیوهیدروژناسیون شکمبه ای اسیدهای چرب غیر اشباع خوراک نسبتاً ثابت است (به ترتیب % 86 و % 82 درصد برای اسید لینولئیک و اسید لینولنیک) (Jenkins and Bridges, 2007). در حالیکه ترشح آنها در شیر بسیار متغیر است (30 تا 60 درصد جذب شده) (Moate et al, 2008) تنظیم مقادیر جذب شده ی اسیدهای چرب که به شیر ترشح می شوند ناشناخته و غیر قابل کنترل است با این حال مقادیر مصرفی با مدیریت کنترل می شوند.
بنابراین می بایستی خوراک حاوی مقادیر زیادی اسیدهای چرب غیر اشباع باشد تا به جذب بیشتر نائل شویم. فرض شده است که 50 درصد اسیدهای چرب ضروری جذب شده به داخل شیر ترشح میشود. پس نیم دیگری از اسیدهای چرب ضروری جذب شده برای اعمال اکسیداسیون و عملکردهای فیزیولوژیک در دسترس می باشند.
بهتر است گاوهای با تولید بالا با مکمل های چربی مثل پنبه دانه، سویا یا پیه تغذیه شوند، این مکملها اسید لینولئیک در دسترس بالایی دارند در حالیکه نشان داده شده که اسیدهای چرب ضروری در دسترس در سویای برشته شده ناچیز است.
علوفه تازه یا تخم بزرک منابع غنی از اسید لینولنیک می باشند. منابع حاوی اسیدهای چرب EPA و DHA روغن ماهی یا جلبک دریایی هستند. پیشنهاد شده است که گاوهای شیرده مقادیر کافی از اسیدهای چرب ضروری را برای حمایت از عملکرد طبیعی فیزیولوژیکی می بایستی مصرف کنند. در همین ارتباط تحقیقات در انسان پیشنهاد می کند که مصرف مقادیر بیشتر اسیدهای چرب n-3 خصوصاً روغن ماهی وضعیت سلامت را در طول دوره های استرس فیزیولوژیک شامل سرطان، بیماری قلبی، بیماری های سیستم ایمنی و جراحی ها بهبود می بخشند.
در مطالعه ای روی گاوهایی با یک و چندشکم زایش با جیره هایی با چربی پایین (کمتر از 2 درصد اسید چرب) یا مکمل شده با 1/7 درصد اسیدهای چرب اشباع شده (Energy Booster 100®) یا 1/7 درصد اسیدهای چرب غیر اشباع (®Megalac) به مدت 60 روز قبل از زایش و به طور مداوم به مدت 90 روز پس از زایش تغذیه شدند. جیره های حاوی چربی غیر اشباع مصرف ماده خشک را هم در گاوهای یک شکم زایش و هم چند شکم زایش قبل از زایمان و تنها ماده گاوهای مسنتر بعد از زایمان را کاهش داد. تعادل انرژی (EB) بعد زایش در هر دو گروه گاوها با تغذیه ی اسیدهای چرب اشباع بهبود یافت. از آنجاییکه مکمل های چربی سبب اختلاف در تولید شیر و درصد چربی شیر می شوند، هیچ اختلافاتی در انرژی تصحیح شده شیر بین تیمارها رخ نداد. در بین معیار های تولید مثلی، اطلاعات اولیه نشان داد که مکمل های چربی دمای ناحیه رکتوم و عفونت های رحمی را کاهش می دهند.
تولید مثل
در بسیاری از مواقع چربی های غذایی بر وضعیت تولید مثلی گاوهای شیری تأثیر می گذارند که شامل افزایش تعداد و اندازه فولیکول های تخمک گذار، افزایش غلظت پروژسترون پلاسما و کاهش ترشح متابولیت پروستاگلاندین می شوند، که سبب افزایش طول عمر جسم زرد و بهبود باروری می شوند (Staples et al., 1998). در حال حاضر اطلاعات کافی به منظور تعیین احتیاجات مصرفی برای بهبود تولید مثل در دسترس نمیباشد.
در ارتباط با تغذیه اسیدهای چرب ضروری به گاوهای شیری دو رویکرد اساسی در نظر گرفته می-شود:
- تغذیه مقادیر زیاد چربی در دوره خشکی یا اوایل شیردهی یا هر دو به منظور اینکه کل لیپیدهای خون افزایش یابند. چون حامل اصلی لیپیدهای خون LDL و HDL می باشند. لیپیدهای بالاتر خون به میزان زیادی فسفولیپیدها و استرهای کلستریل را افزایش داده که اینها خود از اسیدهای چرب غیر اشباع و کلسترول غنی هستند.
- تغذیه منابع غذایی حاوی اسیدهای چرب غیر اشباع لینولئیک و لینولنیک اسید، یا اسیدهای چرب معمولتر 20 و 22 کربنه ی روغن ماهی به ترتیب EPA و DHA در نظر گرفته شوند. غلظتهای بالاتر کلسترول، به عنوان پیش ساز سنتز پروژسترون، سنتز کافی از این هورمون تنظیمی کلیدی را تضمین میکند. هر چند که تحقیقات نشان می دهند که افزایش غلظت لیپید خون سبب افزایش گردش پروژسترون از طریق کاهش پاکسازی (کلیرانس) آن از خون می شود.
تحقیقات نشان داده که در گاوهای شیری پر تولید، تولید شیر بالا به کاهش پتانسیل توسعه ی اووسیت ها در تخمدان می انجامد که با تغذیه مکمل های چربی (800 گرم در روز) منجر به بهبود معنی دار پتانسیل رشد اووسیت می شود. تغذیه مناسب اسیدهای چرب 6- ω و 3-ω تعادل پروستاگلاندین های سری 2 و 3 را تنظیم می کند. تداوم بارداری پس از لقاح نیازمند ترشح پروژسترون مترشحه از جسم زرد (CL) می باشد غلظت های بالای پروستاگلاندین F2α منجر به پس روی جسم زرد شده و نطفه قبل از جایگزینی در رحم از بین می رود. با افزایش در دسترس بودن اسیدهای لینولئیک و لینولنیک سنتز آراشیدونیک اسید کاهش مییابد که خود پیش ساز PGF2α بوده و در نتیجه سنتز آن کاهش می یابد. همچنین مقادیر بالاتر اسید لینولنیک سنتز EPA را افزایش داده که یک پیش ساز پروستاگلاندین های سری 3 بوده که خود اثری مخالف با اثرات PGF2α دارد.
جیرههای حاوی چربیهای غیر اشباع به طور واضحی سهم اسیدهای چرب n-3 و n-6 را در سلول های گرانولوزا و اووسیت ها تحت تأثیر قرار داده و به طور متناظر سبب تغییر نسبت n-6 / n-3 شد، که این خود توانست سهم پروستاگلاندین ها را در این بافت ها تحت تأثیر قرار دهد. بیلبی و همکاران (2006) پروفیل های اسید چرب بافت های متعدد را در گاوهای تغذیه شده با جیره های حاوی پنبه دانه یا نمک های کلسیمی غنی شده با اسیدهای چرب روغن ماهی گزارش کردند.سهم EPA و DHA در اندومتریوم گاوهای تغذیه شده با روغن ماهی افزایش یافت در حالی که سهم آراشیدونیک اسید کاهش یافت. چنین اثراتی برای حفظ کمتر PGF2α و دیگر محصولات پروستاگلاندین سری 2 مثبت خواهد بود، که جسم زرد را حفظ خواهد کرد و غلظت پروژسترون را افزایش داده که احتمالاً منجر به بهبود جایگزینی جنین می گردد.
بایستی به این نکته توجه داشت که اگر مکمل اسید لینولئیک قبل از زایمان بیش از اندازه باشد، فرآیند غیر اشباع سازی و طویل شدن زنجیره آراشیدونیک اسید به عنوان پیش ساز PGF2α محدود خواهد شد. مفهوم دستکاری بیشتر در دسترس بودن اسیدهای چرب ضروری در جهت مدیریت تولید مثل این است که منبعی با اسید چرب لینولئیک اسید از 28 روز قبل زایش تا 28 روز پس از زایش تغذیه کرد، با این حال توصیه می گردد منبعی با اسید چرب n-3 بالا ، ترجیحاً روغن ماهی، از 28 تا 100روز شیردهی دنبال شود. در حالیکه به دنبال استفاده از یک منبع n-3 بالا هستیم می توان پروستاگلاندین های سری 2 را کاهش و سری 3 را در جهت بهبود محیط برای جایگزینی و بقای جنین افزایش داد.
در مجموع این مطالعات پیشنهاد می کند که اگر تغذیه چربی های غنی از اسیدهای چرب غیر اشباع انتخابی، در دوره خشکی شروع و در دوره پس از زایش ادامه یابد، سلامت پس از زایش و تولید شیر و همچنین رشد و نمو جنین گاو و متعاقب آن نرخ آبستنی بهبود می یابد. اثرات سودمند بر روی پاسخ های تولیدمثلی می تواند ناشی از بازیابی سریع سیستم تولید مثلی برای حمایت از نمو جنین باشد.
اصلاح پاسخ ایمنی
سپتی سمی (عفونت خونی) در یک چهارم تا یک سوم گوساله هایی با اسهال شدید رخ داده و بطوری که بیش از نیمی از آنها قادر به ادامه زندگی نخواهند بود. این حالت به واکنش بیش از حد تهاجمی سیستمیک فاز حاد اطلاق می شود که با یک دوره التهاب بیش از حد شناخته می شود و در ادامه از کار افتادگی سیستم ایمنی رخ داده که در پی واکنش ضد التهابی شدید ایجاد می گردد (Ballou et al., 2009). گزارش شده که تغذیهی روغن ماهی موجب کاهش واکنش فاز حاد شده و بقاء حیوان را افزایش می دهد. تصور بر این است که مکمل روغن ماهی می تواند منجر به تعادل بیشتر واکنش فاز حاد با تزریق اندوتوکسین LPS در گوساله های تیپ شیری گردد. تزریق LPS سبب افزایش قابل توجه نرخ تنفس گردید. همه اثرات و بسیاری از جنبه های پاسخ فاز حاد با اضافه کردن روغن ماهی به جایگزین شیر کاهش یافت به طوریکه این اثرات در طیفی بین 5 تا 10 درصد چربی جایگزین شده به صورت اسیدهای چرب روغن ماهی، خطی بودند. بسیاری از اثرات ممکن است با میانجی گری اسید چرب روغن ماهی در تعدیل غلظت پروستاگلاندین بافت مربوط باشند.
دیگر اثرات
به دلیل اینکه بهره وری با مصرف بالاتر اسیدهای چرب 3-ω افزایش می یابد تمایل به مصرف آنها در غذاهای انسانی نیز وجود دارد. رویکردهای گوناگونی نسبت به تحقق این امر در محصولات دامی اخیراً بررسی شده است. علاوه بر مکمل نمودن جیره های دامی با منابع سنتی دارای اسید چرب 3-ω بالا، محصولات جدیدی از طریق مهندسی ژن گیاهی در دسترس میباشند (Whelan, 2009). در برخی گیاهان (مثل سویاها) ژنهایی شناسایی شده اند که میزان اسید استئاریدونیک (3-ω 4 : 18) را افزایش می دهند. زمانی که اسید استئاریدونیک پیش ساز برای طویل سازی زنجیره به ایکوزانوئیدها باشد، محدود کردن آنزیم ∆-6- دسچوراز بی اثر میشود. این اسید چرب به عنوان پیش ساز برای سنتز EPA نسبت به اسید لینولئیک چهار برابرمؤثرتر است.
خلاصه
اسیدهای لینولئیک و لینولنیک برای حیات تمامی پستانداران ضروری می باشند و عملکرد آنها به عنوان اجزاء سازنده غشاء سلولی و همچنین به عنوان پیش ساز برای سنتز پروستاگلاندین ها یا دیگر اسیدهای چرب بلند زنجیر بوده که نقش اساسی را در ساختمان غشاء ایفا می کنند. بنابراین اسیدهای چرب ضروری هم برای ساختار و هم تنظیمات سلولی سلول ضروری هستند. هیچگونه اندازه گیری های مستقیمی در تعیین احتیاجات اسیدهای چرب ضروری نشخوارکنندگان وجود ندارد. به هر حال تحقیقات و مشاهدات پیشنهاد میکنند که جیرههای معمول مقادیر کافی برای عملکرد طبیعی را تأمین میکنند. هیچ گونه اطلاعات جزئی در ارتباط با متابولیسم لینولنیک اسید در نشخوارکنندگان وجود ندارد. چنین تحقیقاتی اگر اثبات گردد می تواند مثمر ثمر باشد. شواهد نشان می دهند که اسیدهای چرب ضروری قادرند به طور مثبتی بر سیستم تولید مثلی و ایمنی مؤثر باشند. تحقیقات همچنان بر اصلاح مدیریت تغذیه اسیدهای چرب ضروری به منظور دسترسی به بهبود عملکرد تولید مثلی می باشد.